
Intracellular compartmentation
Ishier Raote
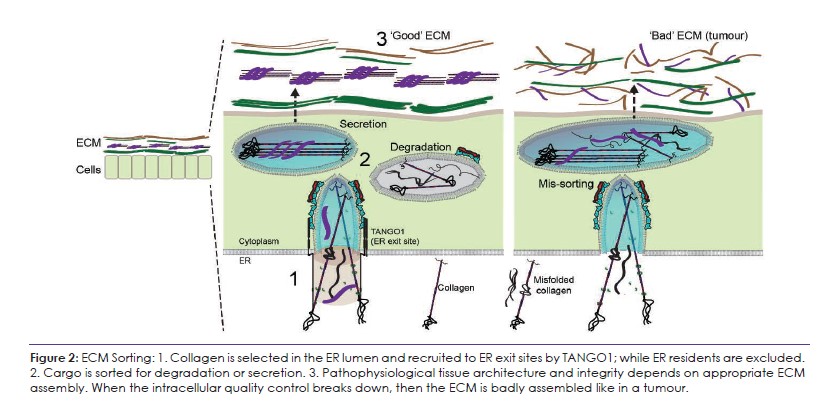
All animals have an extracellular matrix (ECM) – secreted materials that assemble into a biomechanical, 3D scaffold that defines the form of multicellular tissue. The ECM comprises up to 70% of our dry weight and its most abundant components are collagens, which alone comprise 25% of the body’s protein weight. For correct ECM assembly, cells must secrete folded collagens and degrade misfolded collagens. When secretion or degradation go wrong, ECM is misassembled. It remains unclear how cells control the quantity and quality of secreted collagens and other ECM proteins. The Raote lab aims to reveal fundamental mechanisms of collagen sorting in the early secretory pathway and how errors in this control lead to pathophysiological changes.
Mots-clés : extracellular matrix, collagen
All animals have an extracellular matrix (ECM) and its most abundant components are collagens, which alone comprise 25% of the body’s protein weight. For correct ECM assembly, cells must secrete folded collagens while degrading excess or misfolded collagens. When either secretion or degradation go wrong, the resultant ECM is mis-assembled, leading to problems in tissue building and forming aberrant structures such as tumours. Cellular mechanisms of this sorting of intracellular collagens for secretion or degradation in the secretory pathway, remain unknown.
Collagens are a challenging secretory cargo for cells because they are huge. My discoveries of TANGO1 as a master organiser of the ERES-Golgi interface, have provided mechanistic insight into how bulky and high-volume cargoes like collagen are secreted. These studies provide an ideal platform to reveal fundamental mechanisms of how collagen is sorted between secretion and degradation, and how the process of sorting contributes to tissue building.
The early secretory pathway, at the interface between endoplasmic reticulum (ER) exit sites (ERES), the ER-Golgi Intermediate Compartment (ERGIC), and the Golgi, is a vital gateway to control the quantity and quality of collagens, which then go on to assemble into an ECM. My group aims to reveal mechanisms how TANGO1 and its associated ERES machinery act as a collagen quality and quantity control checkpoint, providing insights into the pathophysiological conditions associated with altered ECM and tissue assembly including tumorigenesis (Figure 2). To start, we shall continue to use collagen as a model, subsequently including other large ECM proteins including laminin, fibrillin 1, and fibronectin. Our studies of intracellular steps in collagen secretion, we will:
1. Reconstitute collagen ‘filtration’ at the ERES, to understand how unfolded (premature) collagens are prevented from exiting the ER
When collagen folds, it forms a rigid triple helical rodlike structure. This stability and rigidity is vital for correct ECM assembly. Unfolded (premature) collagen should not be allowed to leave the ER and be secreted. It is seen that premature proteins are retained at ERES and prevented from escaping, but the mechanism for this active ER retention remains unknown. Our preliminary evidence suggests that TANGO1 plays a role in this process and in particular its extensive intrinsically disordered regions (IDRs) in the ER lumen. We aim to study how the IDRs contribute to this kind of collagen ‘filtration’ in the ER lumen.
One hypothesis suggests that TANGO1 IDRs undergo liquid-liquid phase separation, forming a specialised sub-compartment within the ER lumen, which separates folded collagen from premature collagen and other ER residents that should not escape from the ER.
Our group will use a bottom up approach with DNA origami nanopores to reconstitute the dynamics and physical state of IDRs lining the pore under different conditions. In a top down approach, we will generate targeted mutations in TANGO1 and related ERES machinery to identify other components of the filter and how they all work together.
2. Sorting of terminally misfolded collagens for degradation at an ERES
An unusual problem during collagen folding is that sometimes, instead of forming a correctly folded rod-like rigid triple helix, there are interruptions in the rod caused by local errors in folding. These misfolded collagens now resemble a rod with a ‘hinge.’ These misfolded collagens cannot be allowed to be secreted and must be degraded, but the mechanisms for their degradation are poorly understood.
Recent reports show that the ERES is not only a site to export secretory cargoes, but even to degrade excess, or terminally misfolded collagen, by routing it to lysosomes instead of toward the Golgi. There is almost no understanding at all of how ERES machinery is utilised in this sorting. We will identify this molecular machinery and how it is repurposed for the two in secretory or degradative trafficking pathways. We will knock-in ‘BioID’ tags at the cytoplasmic terminal of TANGO1 in fibroblasts and keratinocytes (CCL1106 cells) to biotinylate proteins in the proximity of the ERES under conditions of secretion or degradation. Biotinylated proteins will be immunoprecipitated and identified by mass spectrometry. Target genes will be knocked out using CRISPR/Cas9 to test their role in collagen quality and quantity control in fibroblasts and keratinocytes.
3. Pathophysiological alterations in in skin tumours that arise when ERES sorting fails. With collaborator Dr Neil Rajan (Newcastle, UK)
We collaborate with Dr Neil Rajan, a clinical dermatogeneticist, who is an expert in rare genetic skin disorders. Together we have identified a rare disorder, CYLD cutaneous syndrome (CCS) with large cutaneous tumours, where ERES machinery (particularly TANGO1) is degraded. Since TANGO1 is required for collagen secretion, one would imagsine that tumours secrete less collagen – but counterintuitively, tumours hypersecrete collagen! We discovered that secreted collagen is often misfolded, even though it should never have left the ER, reinforcing the role of TANGO1 and other ERES machinery as a quality and quantity control checkpoint. We will study ECM mis-assembly in these skin tumours, focusing on the effects of altered quality control on ECM. Tumour cells derived from CCS patients will be used to characterise differences in tumour cell ERES and identify how the ERES checkpoint is lost in tumours. One focus will be on revealing the mechanisms through which tumours degrade TANGO1.
Overall, this research on collagen filtration, sorting and degradation has important implications for understanding and treating tumorigenesis or ECM hypersecretion disorders like fibrosis, which contribute to ~45% of all deaths in industrialised nations.

Pour contacter un membre de l’équipe par mail : prenom.nom@ijm.fr
Publications
Preprint
Review
Fondation ARC, FRM